En général, le
système respiratoire est l'ensemble des organes qui assurent l'apport d'oxygène
et l'élimination du gaz carbonique. On lui distingue une partie conductrice et
une partie purement respiratoire, les alvéoles pulmonaires. La partie
conductrice est une succession de tubes qui relient les alvéoles au milieu
extérieur. Elle est subdivisée en voies aériennes supérieures et profondes; les
voies aériennes profondes sont extra- ou intra pulmonaires.
L'origine du système respiratoire est
essentiellement endoblastique; seules, les fosses nasales sont ectoblastiques.
Il débute sous forme d'une évagination de la face ventrale du pharynx. Cette
évagination, représentée en bleu et située en arrière des poches pharyngiennes
(I à IV), s'isole du tube digestif primitif tout en maintenant sa communication
avec le pharynx et se ramifie progressivement. A la naissance, les premières
inspirations vont déployer son extrémité distale et ouvrir les alvéoles.
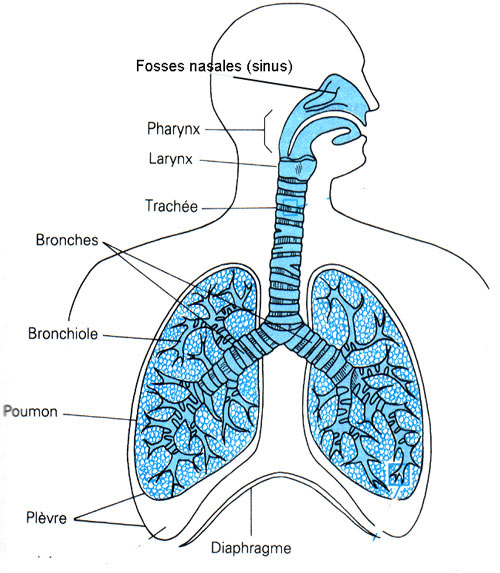
Ce système respiratoire comprend les voies aériennes supérieures.
A) Les voies aériennes supérieures
Les voies aériennes supérieures comprennent
les fosses nasales, le pharynx subdivisé en nasopharynx , oropharynx et laryngopharynx , et enfin le larynx .
 |
| Le système respiratoire |
a) Fosses
nasales
1).
Microanatomie
Dans cette coupe sagittale de la tête, la
paroi latérale d'une fosse nasale et ses cavités osseuses annexes, les sinus,
sont dessinés en orange. Trois expansions osseuses latérales y forment les
cornets. Les cornets inférieur et moyen sont importants et partiellement
sectionnés. Le cornet supérieur est un petit repli au-dessus du cornet moyen.
La fosse nasale
est divisée en trois parties. La partie antérieure ou vestibulaire, très petite,
est tapissée de vibrisses et correspond aux narines dont la paroi est
essentiellement cartilagineuse. La partie postérieure ou respiratoire comprend
presque toute la cavité, donc les cornets inférieur et moyen. Elle communique
par de fins pertuis avec les sinus; seuls le sinus frontal et le sinus
sphénoïdal sont représentés.
La partie
supérieure des fosses nasales ou région olfactive est plus limitée. Elle
s'étend du cornet supérieur à la voûte des fosses nasales et recouvre la partie
supérieure de la cloison médiane. Les faisceaux de fibres nerveuses olfactives
traversent les orifices de la lame criblée de l'ethmoïde pour rejoindre le
bulbe olfactif.
Cette coupe
oblique de fosses nasales, chez un embryon de rat, permet d'observer les
régions vestibulaire, respiratoire et olfactive. Leur localisation n'est pas
identique à ce que l'on observe dans l'espèce humaine mais leur structure
histologique est comparable.
La cloison
médiane est faite de deux parties. La partie arrière, en haut dans l'image, est
un cartilage dont l'extrémité antérieure est renflée et dont l'extrémité postérieure
est bifide. Devant ce cartilage se trouve un massif osseux qui contient deux
petites cavités, les organes olfactifs. De part et d'autre de la cloison
médiane il existe une cavité allongée et irrégulière, la fosse nasale
proprement dite. Elle est incomplètement cloisonnée par les cornets, replis
osseux recouverts par une muqueuse. La paroi externe est osseuse et colorée en
bleu. En arrière, les fosses nasales sont limitées par deux os plats;
latéralement elles sont limitées par le massif spongieux du maxillaire
supérieur. Celui-ci contient une volumineuse ébauche dentaire et, entre cette
ébauche et la fosse nasale, la cavité du sinus maxillaire. Le maxillaire est
recouvert par quelques faisceaux de muscle squelettique. Les fosses nasales se
prolongent vers l'avant dans le vestibule nasal dont l'épithélium est
épidermoïde.
2). Muqueuse
nasale
La muqueuse qui
tapisse plus des 2/3 des fosses nasales est limitée par un épithélium
pseudostratifié de type respiratoire. Il est composé de cellules cylindriques
ciliées, de cellules caliciformes non visibles dans la zone choisie et de
petites cellules basales ou cellules de réserve. La pseudostratification et
encore peu marquée dans cette préparation d'embryon. Les petites cellules
basales qui permettent la régénération épithéliale sont à ce point nombreux
qu'elles forment pratiquement une assise continue.
Le chorion est
mésenchymateux dans cette coupe embryonnaire mais lorsqu'il est bien
différencié, il est relativement dense et riche en fibres élastiques. Des
glandes tubuleuses séreuses, muqueuses ou mixtes déversent leurs sécrétions
dans une petite invagination épithéliale toujours ciliée. Ces sécrétions
glandulaires humidifient la cavité nasale. Leur contrôle est autonomique,
contrairement à celui des cellules caliciformes dont la stimulation dépend de
la température, de l'humidité et de la contamination de l'air inhalé.
La
vascularisation de la muqueuse respiratoire est particulière puisqu'elle
comprend des shunts artérioveineux et un plexus veineux caverneux,
particulièrement développé au niveau des cornets.
Ce faible
grossissement de cornets, prélevés après fixation par perfusion, démontre leur
importance et surtout celle de leur plexus veineux dilaté. Le squelette osseux
des cornets est mince, ramifié et intensément coloré. La muqueuse, limitée en
dehors par la bande rouge de l'épithélium, est remplie de cavités qui sont les
veines dilatées vidées de leur sang par la perfusion.
L'engorgement du
plexus, contrôlé par les shunts artérioveineux, est responsable de la
turgescence de la muqueuse respiratoire qui peut parfois obstruer la fosse
nasale. Cette turgescence s'accompagne d'un transsudat et permet une meilleure
humidification de l'air inhalé. Chez tout individu dont la fonction nasale est
normale, les veines s'ouvrent et se ferment périodiquement et ces réactions
vasomotrices alternent d'une fosse nasale à l'autre. Ce cycle nasal dure
environ 3 heures. Les phénomènes vasculaires nasaux sont complexes et réglés
par les voies nerveuses ortho- et parasympathiques mais aussi par les
propriétés de l'air.
Puisqu'elles
forment le début des voies respiratoires, les fosses nasales sont les plus
sujettes aux agressions répétées. Il est donc normal d'y trouver de nombreuses
lésions épithéliales : à certains endroits, l'épithélium respiratoire est
démuni de cils, devient cylindrique, simple puis cubique, ou même cubique
aplati. Ailleurs, l'épithélium peut devenir épidermoïde.
La muqueuse
nasale est couverte d'un film de mucus, formé en grande partie d'eau (95%) mais
aussi de glycosaminoglycanes, de glycoprotéines, de protéines et d'ions. Le
mucigène est produit par les cellules caliciformes et surtout par les glandes
muqueuses et mixtes du chorion. La phase aqueuse est assurée par une
transsudation qui dépend d'un gradient de pression osmotique et par la
sécrétion des glandes séreuses et mixtes.
A la surface
cellulaire, le tapis muqueux est organisé en deux couches. La couche
superficielle repose sur l'extrémité des cils; elle est visqueuse, parce que
riche en glycosaminoglycanes. La couche profonde est beaucoup plus fluide et
permet le battement ciliaire. Sous l'action des cils, la couche superficielle
glisse en direction du pharynx, en entraînant les grosses poussières de l'air
inhalé.
3). Organe
olfactif
L'organe
olfactif de ce rongeur est limité par deux épithéliums pseudostratifiés
différents. L'un est mince et de type respiratoire; l'autre, beaucoup plus
épais, est olfactif. L'étendue de la zone olfactive et la transition entre les
deux épithéliums diffèrent selon les espèces.
Dans
l'épithélium olfactif, les noyaux de certaines cellules sont superficiels et
allongés; leur disposition régulière forme une ligne dense à la surface de
l'épithélium. Les noyaux des autres cellules sont profonds, et leur disposition
est irrégulière.
L'organisation
cellulaire de l'épithélium pseudostratifié olfactif est plus nette ici bien que
la pseudostratification n'y soit pas encore évidente. Les cellules dont les
noyaux sont les plus superficiels sont des cellules épithéliales vraies,
appelées cellules de soutien; les cellules plus profondes sont des cellules
nerveuses bipolaires olfactives qui ont colonisé l'épithélium. Un axone d'une
cellule bipolaire est désigné à gauche par une flèche et une dendrite à droite
par une tête de flèche.
Les cellules de
soutien, claires dans ce schéma sont plus massives que les cellules bipolaires
bleues, ont leur noyau dans le tiers supérieur de l'épithélium. Leur pôle
apical est pourvu de microvillosités assez longues et renferme un important réticulum
endoplasmique lisse tandis que leur pôle basal est souvent effilé et s'insinue
entre les cellules voisines. Leurs microfilaments, mitochondries, lysosomes et
corps résiduels, représentés en jaune, sont abondants. Quelques petites
cellules basales non différenciées permettent leur remplacement.
Les cellules
olfactives ont un corps cellulaire situé à des niveaux variables et sont
responsables de l'aspect pseudostratifié de l'épithélium. Elles sont unies aux
cellules de soutien par des complexes de jonction. Leur expansion dendritique
apicale se termine par la vésicule olfactive qui aboutit dans le mucus et se
prolonge par une dizaine de cils immobiles, parallèles à la surface
épithéliale. La dendrite et sa vésicule sont largement pourvues en mitochondries,
microtubules et neurofilaments. Ces derniers se retrouvent aussi dans le corps
cellulaire et dans l'axone.
Les axones de
cellules olfactives voisines traversent la mince couche basale épithéliale,
sont enveloppés par des cellules de Schwann et se réunissent dans le chorion ou
parfois déjà dans l'épithélium pour former des faisceaux nerveux amyélinisés,
les filets olfactifs. Ils cheminent vers les pertuis de la lame criblée de
l'ethmoïde.
D'autres
dendrites myélinisées, représentées en noir, appartiennent au nerf trijumeau.
Leur extrémité non myélinisée se trouve dans l'épithélium et forme à sa surface
de fines arborisations, qui ne sont pas dessinées. Elles seraient réceptrices
de stimuli non olfactifs.
Il existe sous
l'épithélium de petites glandes acineuses mixtes, les glandes de Bowman. Leur
richesse en lipofuscines, ainsi que celle des cellules de soutien, serait
responsable de la couleur jaune de la région olfactive.
L'irrégularité de
la surface olfactive est démontrée par la microscopie à balayage. Les
microvillosités (MV) des cellules de soutien s'étendent entre les vésicules
olfactives (OV) dont la hauteur varie en fonction de la perception olfactive.
C'est pourquoi la vésicule désignée par un astérisque est enfoncée dans
l'épithélium, celle désignée par une flèche ne l'est qu'à moitié. A certains
endroits, les cils (Ci) sont modifiés (MC) et deviennent aussi minces que les
microvillosités des cellules de soutien. Le rôle des cellules de soutien n'est
pas connu mais il paraît un peu simpliste de leur accorder uniquement une
fonction de support.
Les glandes de
Bowman déversent leur sécrétion à la limite entre les deux épithéliums
pseudostratifiés. Elles sont souvent décrites comme mixtes, composées de deux
types cellulaires. Dans cette préparation, toutes les cellules semblent
identiques. Ces glandes sont comparables aux glandes de von Ebner associées aux
papilles circonvallariées. Elles humidifient et nettoient la muqueuse, mais interviennent
aussi dans l'olfaction en permettant la dissolution chimique des substances
odorantes.
L'histophysiologie
de l'olfaction est encore mal connue. Il y a 2000 ans, Lucrèce, un poète latin,
proposa une explication très simple de l'odorat. Il imaginait que le palais
contenait de minuscules pores de forme et de taille différente et que les
substances odorantes dégageaient de petites particules qui s'emboîtaient
exactement dans un port qui leur était destiné et qui permettait
l'identification de chaque odeur.
L'idée de
Lucrèce était exacte. La plupart des odeurs sont déterminées par la géométrie
et la taille des molécules odorantes. Parmi les composés odorants, on a pu
sélectionner sept odeurs primaires. Ce sont le camphre, le musc, la fleur, le
menthol, l'éther, l'odeur piquante et l'odeur putride. Pour chaque odeur
primaire, il existerait un récepteur sur la membrane de l'extrémité dendritique
de la cellule bipolaire, chaque cellule bipolaire n'ayant sans doute qu'un type
de récepteur. Un récepteur ne peut accepter que la molécule dont la
configuration lui est appropriée bien que certaines molécules s'adaptent
probablement à plusieurs types de cavités. La liaison de la molécule avec sa
cavité réceptrice engendre des potentiels d'action qui sont véhiculés par les
axones vers le bulbe olfactif et de là aux centres supérieurs du cerveau.
Résumons les
fonctions des fosses nasales. Grâce à leur architecture, à leur vascularisation
importante et à leur activité sécrétrice, elles permettent non seulement la conduction
de l'air mais aussi son humidification et son réchauffement.
En plus de leur
fonction olfactive, les fosses nasales, grâce à leurs vibrisses et à leur
appareil mucociliaire éliminent les grosses particules mélangées à l'air
inspiré.
b) Pharynx
Le pharynx appartient en partie au tube
digestif. On décrit à sa paroi une muqueuse, une sous-muqueuse inconstante et
une musculeuse.
La muqueuse est
délimitée suivant les endroits par un épithélium respiratoire ou épidermoïde.
Le tissu conjonctif sous-épithélial est dense et riche en fibre élastique. La
sous-muqueuse est un tissu conjonctif dense.
La musculeuse,
faite des muscles striés, est organisée en une couche interne longitudinale et
une couche externe oblique ou circulaire. Entre les faisceaux musculaires, il
existe des nombreuses fibres élastiques.
Le pharynx est
le carrefour des voies respiratoires et digestives. Dans ses trois parties,
naso , oro et laryngopharynx ,
l'épithélium de sa muqueuse est différent.
Le nasopharynx
fait suite aux fosses nasales et est comme elles, tapissé par un épithélium
pseudostratifié de type respiratoire. Il renferme dans sa paroi dorsale
supérieure des formations lymphoïdes constituant l'amygdale pharyngée et dans
sa paroi latérale, les amygdales tubaires. L'amygdale pharyngée, marquée par
des replis irréguliers, est bien développée chez le nouveau-né mais s'atrophie
après la puberté. Les amygdales tubaires sont localisées près de l'orifice de
la trompe d'Eustache. Elles ne sont pas représentées mais le sillon schématisé
dans la cavité pharyngienne indique approximativement leur localisation.
L'oropharynx
prolonge la cavité buccale et, comme elle, est revêtu d'un épithélium
pluristratifié épidermoïde. Le même épithélium se retrouve dans le
laryngopharynx et l'oesophage.
c) Larynx
Le larynx fait partie des voies respiratoires
conductrices supérieures et est aussi le siège de la phonation. Il est formé
d'un squelette cartilagineux coloré en bleu-gris dont les pièces sont reliées
par des muscles striés colorés en rouge et par des ligaments colorés en orange.
Seuls les cartilages impairs sont représentés. L'épiglottique est de type
élastique et ne s'ossifie jamais. Les cartilages thyroïde et cricoïde sont tous
deux de type hyalin et s'ossifient avec l'âge. Le premier anneau trachéen et
les deux lobes thyroïdiens sont également représentés.
La cavité
laryngée est limitée par une muqueuse de type respiratoire. Elle est
irrégulière et formée d'une région supérieure ou vestibulaire, de deux
ventricules latéraux et d'une cavité infraglottique. Les quatre replis de la
muqueuse qui forment les ventricules latéraux sont en haut les fausses cordes
vocales et en bas les vraies cordes vocales. Les vraies cordes vocales sont
recouvertes d'un épithélium pluristratifié épidermoïde, dessiné en vert, et
délimitent un espace de taille variable, la glotte. Le chorion de cet
épithélium est dépourvu de glandes. Il renferme une large bande de tissu
élastique, le ligament vocal, bordé par le muscle vocal ou thyro-aryténoïdien.
La contraction des muscles vocaux modifie l'espace entre les cordes vocales, ce
qui change la hauteur des sons au passage de l'air.
Voici la coupe
du larynx d'un jeune chien à l'endroit où le carrefour aéro-digestif se divise.
Les cavités laryngée et œsophagienne ne sont pas encore tout à fait
individualisées et sont limitées par le même épithélium épidermoïde. Les masses
musculaires et cartilagineuses sont à ce point développées qu'elles forment la
majeure partie de la paroi. Quelques glandes mixtes, plus nombreuses à la
jonction des deux organes, s'étendent dans le chorion mince du larynx.
Les glandes
laryngées, comme celles de la trachée et des bronches, sont mixtes et
habituellement classées parmi les glandes tubuleuses car leur forme est
grossièrement celle d'un tube. On distingue toutefois des tubes muqueux et
séreux. Leur association est très variable. Parfois ils aboutissent séparément
dans une dilatation collectrice; à d'autres endroits, comme ici, la partie
séreuse d'un tube est poursuivie par une partie muqueuse qui se jette dans la
dilatation. Celle-ci peut elle-même être sécrétrice de mucus ou simplement
excrétrice. L'importance des deux parties sécrétrices et de la partie
collectrice varie suivant leur localisation. Dans chaque glande, les cellules
séreuses et muqueuses sont entourées par des cellules myoépithéliales, non
visibles à ce grossissement.
B) Les voies
aériennes profondes
Les voies
aériennes profondes extra pulmonaires comportent la trachée et les deux bronches souches. Celles-ci
pénètrent dans les poumons au niveau du hile. Les voies profondes intra
pulmonaires se divisent dans le poumon droit en trois bronches lobaires et, dans
le poumon gauche, en deux bronches lobaires. A leur tour, celles-ci se
ramifient en 10 bronches segmentaires à droite et 8 à gauche. Les bronches
segmentaires se subdivisent ensuite en bronches de plus en plus petites qui ne
sont pas représentées dans ce schéma et qui se terminent par des bronchioles.
1) Trachée
La trachée, comme les bronches souches, fait
partie des voies conductrices profondes extrapulmonaires. Chez l'homme, c'est
un tube flexible de 11 cm de long et 2 cm de large formé de 16 à 20 anneaux
cartilagineux incomplets superposés. Cette coupe transversale est oblique et
passe par deux anneaux successifs, leur forme typique en fer à cheval
n'apparaît donc pas. En outre, la préparation a rapproché les deux extrémités
du fer à cheval dans la partie dorsale de cette trachée. Cette région dorsale
est repérable parce que la paroi y est plissée. On y trouve un muscle lisse, le
muscle trachéal, qui a l'aspect d'une mince bande violette et qui unit les
extrémités de l'anneau cartilagineux. Dans certaines préparations, il peut être
refoulé dans la muqueuse.
La paroi de la
trachée est relativement simple. Elle comprend une muqueuse, une couche
fibro-cartilagineuse et une couche conjonctive externe. La muqueuse est de type
respiratoire; son épithélium est pseudostratifié et son chorion bien
vascularisé et riche en fibres élastiques renferme quelques glandes mixtes.
La couche
fibro-cartilagineuse comprend les anneaux cartilagineux et leur périchondre
fait de tissu conjonctif dense. Les anneaux cartilagineux sont unis par du
tissu fibreux qui contient de nombreuse fibres élastiques.
La couche
conjonctive la plus externe est riche en tissu adipeux, vaisseaux et nerfs.
La
pseudostratification épithéliale est plus marquée que dans les fosses nasales
parce que les cellules cylindriques sont plus hautes. Entre les cellules
ciliées s'intercalent des cellules caliciformes. Elles peuvent se rassembler
dans de petits replis, les glandes intraépithéliales, que l'on ne voit pas ici.
La membrane basale est très épaisse et ne nécessite pas de coloration spéciale
pour être mise en évidence. Le chorion renferme de nombreuses fibres élastiques
longitudinales. De petites glandes tubuleuses mixtes débouchent dans une
invagination épithéliale ciliée.
La
vascularisation de la trachée dépend surtout des artères thyroïdiennes
inférieures. Un réseau lymphatique (L), situé dans la partie profonde du
chorion, est drainé par les nombreux ganglions péritrachéaux. L'innervation
provient du nerf vague et de la chaîne sympathique.
L'appareil
ciliaire de la paroi trachéale est bien visible au microscope à balayage. Les
cils (Ci) sont estimés à plus de 200 par cellule. Ils battent en direction du
pharynx et font ainsi remonter le mucus à une vitesse de 13 mm par minute. Plus
on s'élève dans l'ascenseur mucociliaire, plus la vitesse du battement ciliaire
ainsi que la proportion de cellules ciliées s'accroissent. Quelques cellules
caliciformes (GC), sont éparpillées dans la paroi.
La trachée a
donc pour fonction, non seulement de conduire l'air et de l'humidifier, mais aussi
de le purifier à l'aide de son important appareil mucociliaire.
2) Bronches
2.1. Bronches
souches
Hors des
poumons, les deux bronches souches ont la même structure que la trachée bien
que leur calibre soit inférieur et que les anneaux cartilagineux soient
complets. Dans les poumons, les anneaux cartilagineux sont fragmentés.
Au fur et à
mesure que les bronches se subdivisent, la structure de leur paroi se modifie.
L'épithélium est encore pseudostratifié dans les bronches lobaires; il devient
cylindrique cilié dans les bronches segmentaires et le reste dans ses
subdivisions; dans les bronchioles dont le diamètre est d'environ 1 mm,
l'épithélium devient cubique puis perd ses cils. Les cellules caliciformes,
encore nombreuses dans les bronches segmentaires, se raréfient progressivement
et n'existent plus dans les dernières bronchioles.
Dans le chorion,
les glandes et les nodules de cartilage diminuent petit à petit et les
bronchioles en sont dépourvues. Par contre les cellules musculaires s'agencent
en une musculeuse incomplète, le muscle de Reissessen, dès les bronches
souches, augmentent relativement dans les bronches segmentaires et persistent
en petit nombre jusque dans les dernières bronchioles.
2.2 Bronches
segmentaires
La bronche segmentaire a une lumière
régulière, bordée par un épithélium cylindrique. Son chorion mince renferme des
nodules cartilagineux de taille et de forme variables. Nous décrirons plus tard
les nombreuses alvéoles et les vaisseaux qui l'entourent.
Son épithélium
est cylindrique simple et cilié; il renferme encore des cellules caliciformes.
Les petits faisceaux musculaires lisses du chorion sont en majorité coupés
transversalement parce que leur disposition est, suivant les endroits, spiralée
ou circulaire. Les fibres conjonctives sont essentiellement élastiques. Les
glandes mixtes et le cartilage sont profonds.
2.3. Bronchioles
Dans la
bronchiole, l'épithélium est cubique simple et le chorion ne renferme plus que
quelques cellules musculaires lisses annulaires et des fibres de collagène. A
côté de la bronchiole se trouve une branche de l'artère pulmonaire.
La paroi de la
bronchiole est mince mais son épithélium comprend des cellules cubiques, dont
certaines ont encore des cils courts, peu visibles, et d'autres, les cellules
de Clara, dont le pôle apical bombe dans la lumière. Chez le rat et la souris,
elles se retrouvent dans tout l'arbre bronchique. Dans l'espèce humaine, elles
sont beaucoup moins nombreuses et n'existent que dans les dernières
bronchioles.
Le seul
caractère morphologique constant de la cellule de Clara est son pôle apical qui
bombe entre les cellules ciliées. Il contient de nombreuses mitochondries
ovoïdes et des granules denses volumineux. Le noyau de la cellule est basal,
souvent irrégulier, et entouré de réticulum endoplasmique rugueux et du système
de Golgi.
Le pôle apical
d'une cellule de Clara fait ici largement sailli dans la lumière. Sa membrane
plasmique est irrégulière, elle présente plusieurs invaginations. Une
évagination est encore plus importante dans une cellule voisine. Les grains ont
des formes et des tailles variables; certains sont fusionnés. Sur la base de
pareilles images, on suppose que la cellule de Clara peut sécréter de trois
façons. Dans les conditions normales, sa sécrétion est mérocine, par exocytose
des grains. Lorsque la sécrétion est stimulée, elle serait apocrine; la cellule
serait alors amputée de son pôle apical ou de ses évaginations. Certains
produits pourraient aussi quitter la cellule par diffusion.
La cellule de
Clara sécrète des lipides et des protéines dont certaines sont des enzymes.
Elle intervient dans la composition du surfactant par l'action de ses
phospholipases. Elle interviendrait dans certaines détoxications grâce à sa
richesse en oxydases liées aux cytochromes P450. Elle pourrait enfin se
transformer en un autre type cellulaire épithélial.
L'arbre
bronchique contient aussi des cellules endocrines qui sont isolées dans son
épithélium ou regroupées en petits amas appelés corps neuro-épithéliaux.
Ceux-ci sont surtout localisés aux différents embranchements de l'arbre
bronchique. Ils contiennent deux types de cellules : les unes sont en contact
direct avec la lumière bronchique par leur pôle apical; les autres par contre,
n'arrivent pas en contact avec celle-ci. Par l'action de leurs sécrétions sur
les muscles lisses vasculaires et bronchiques, ces cellules endocrines participent
à la régulation de la vascularisation et de la ventilation pulmonaire.
2.4. Bronchioles
terminales et respiratoires
La dernière
bronchiole purement conductrice s'appelle bronchiole terminale. Elle est
accompagnée d'une artériole pulmonaire et donne naissance à plusieurs
bronchioles respiratoires, dont un seul est visible ici. Sa paroi est
interrompue par l'évagination des alvéoles (têtes de flèche), que nous verrons
en détail dans la leçon prochaine. Chez l'homme, il existe trois générations de
bronchioles respiratoires. Elles sont courtes et leur diamètre est compris
entre 0,5 et 1 mm. Chaque bronchiole respiratoire se subdivise en une dizaine
de canaux alvéolaires, dont la paroi est fragmentée. Un des fragments est
désigné par une flèche.
Voici une
portion de bronchiole respiratoire. A deux reprises, son épithélium cubique
bas, s'arrête brusquement pour faire place à une alvéole et réapparaît tout
aussi brusquement. Les cellules musculaires lisses sont rares et donc
difficiles à voir.
2.5 Canal
alvéolaire
Voici la
transition entre la bronchiole respiratoire et un canal alvéolaire. La paroi de ce dernier
est limitée à quelques saillies qui sont les bourrelets d'insertion alvéolaire.
Ils sont formés de fibres de collagène ou élastiques et de quelques cellules
musculaires lisses. Suivant les endroits, ils sont recouverts d'un épithélium
cubique aplati ou pavimenteux, en continuité avec l'épithélium alvéolaire. Leur
structure détaillée n'est pas visible dans cette préparation.
Le canal
alvéolaire se termine en cul-de-sac. Son extrémité est dessinée par une ligne
pointillée qui relie les derniers bourrelets d'insertion alvéolaire.
La leçon
suivante sera consacrée aux alvéoles pulmonaires, aux réseaux vasculaires, à
l'innervation des poumons et à la plèvre.